Основные органоиды растительной и животной клетки. Клеточные органоиды: их строение и функции. Жизненный цикл клетки
Органоиды - это постоянные компоненты клетки, выполняющие определенные функции.
В зависимости от особенностей строения их делят на мембранные и немембранные. Мембранные органоиды, в свою очередь, относят к одномембранным (эндоплазматическая сеть, комплекс Гольджи и лизосомы) или двумембранным (митохондрии, пластиды и ядро). Немембранными органоидами являются рибосомы, микротрубочки, микрофиламенты и клеточный центр. Прокариотам из перечисленных органоидов присущи только рибосомы.
Строение и функции ядра. Ядро - крупный двумембранный органоид, лежащий в центре клетки или на ее периферии. Размеры ядра могут колебаться в пределах 3-35 мкм. Форма ядра чаще сферическая или эллипсоидная, однако имеются также палочковидные, веретеновидные, бобовидные, лопастные и даже сегментированные ядра. Некоторые исследователи считают, что форма ядра соответствует форме самой клетки.
Большинство клеток имеет одно ядро, но, например, в клетках печени и сердца Их может быть два, а в ряде нейронов - до 15. Волокна скелетных мышц содержат обычно много ядер, однако они не являются клетками в полном смысле этого слова, поскольку образуются в результате слияния нескольких клеток.
Ядро окружено ядерной оболочкой, а его внутреннее пространство заполнено ядерным соком, или нуклеоплазмой (кариоплазмой ), в которую погружены хроматин и ядрышко. Ядро выполняет такие важнейшие функции, как хранение и передача наследственной информации, а также контроль жизнедеятельности клетки (рис. 2.30).
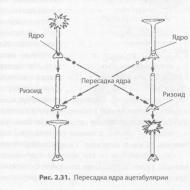
Роль ядра в передаче наследственной информации была убедительно доказана в экспериментах с зеленой водорослью ацетабулярией. В единственной гигантской клетке, достигающей в длину 5 см, различают шляпку, ножку и ризоид. При этом она содержит только одно ядро, расположенное в ризоиде. В 1930-е годы И. Хеммерлинг пересадил ядро одного вида ацетабулярии с зеленой окраской в ризоид другого вида, с коричневой окраской, у которого ядро было удалено (рис. 2.31). Через некоторое время у растения с пересаженным ядром выросла новая шляпка, как у водорос- ли-донора ядра. В то же время отделенные от ризоида шляпка или ножка, не содержащие ядра, через некоторое время погибали.

Ядерная оболочка образована двумя мембранами - наружной и внутренней, между которыми есть пространство. Межмембранное пространство сообщается с полостью шероховатой эндоплаз- матической сети, а наружная мембрана ядра может нести рибосомы. Ядерная оболочка пронизана многочисленными порами, окантованными специальными белками. Через поры происходит транспорт веществ: в ядро попадают необходимые белки (в т. ч. ферменты), ионы, нуклеотиды и другие вещества, и покидают его молекулы РНК, отработанные белки, субъединицы рибосом.
Таким образом, функциями ядерной оболочки являются отделение содержимого ядра от цитоплазмы, а также регуляция обмена веществ между ядром и цитоплазмой.
Нуклеоплазмой называют содержимое ядра, в которое погружены хроматин и ядрышко. Она представляет собой коллоидный раствор, по химическому составу напоминающий цитоплазму. Ферменты нуклеоплазмы катализируют обмен аминокислот, нуклеотидов, белков и др. Нуклео-плазма связана с гиалоплазмой через ядерные поры. Функции нуклеоплазмы, как и гиалоплаз-мы, состоят в обеспечении взаимосвязи всех структурных компонентов ядра и осуществлении ряда ферментных реакций.
Хроматином называют совокупность тонких нитей и гранул, погруженных в нуклеоплазму. Выявить его можно только при окрашивании, так как коэффициенты преломления хроматина и нуклеоплазмы приблизительно одинаковы. Нитчатый компонент хроматина называют эухро-матином, а гранулярный - гетерохроматином. Эухроматин слабо уплотнен, поскольку с него считывается наследственная информация, тогда как более спирализованный гетерохроматин является генетически неактивным.
Хроматин представляет собой структурное видоизменение хромосом в неделящемся ядре. Таким образом, хромосомы постоянно присутствуют в ядре, изменяется лишь их состояние в зависимости от функции, которую ядро выполняет в данный момент.
В состав хроматина в основном входят белки-нуклеопротеины (дезоксирибонуклеопротеины и рибонуклеопротеины), а также ферменты, важнейшие из которых связаны с синтезом нуклеиновых кислот, и некоторые другие вещества.
Функции хроматина состоят, во-первых, в синтезе специфических для данного организма нуклеиновых кислот, которые направляют синтез специфических белков, во-вторых, в передаче наследственных свойств от материнской клетки дочерним, для чего хроматиновые нити в процессе деления упаковываются в хромосомы.
Ядрышко - сферическое, хорошо заметное под микроскопом тельце диаметром 1-3 мкм. Оно формируется на участках хроматина, в которых закодирована информация о структуре рРНК и белках рибосом. Ядрышко в ядре часто одно, однако в тех клетках, где происходят интенсивные процессы жизнедеятельности, ядрышек может быть два и более. Функции ядрышек - синтез рРНК и сборка субъединиц рибосом путем объединения рРНК с белками, поступающими из цитоплазмы.
Митохондрии - двумембранные органоиды округлой, овальной или палочковидной формы, хотя встречаются и спиралевидные (в сперматозоидах). Диаметр митохондрий составляет до 1 мкм, а длина - до 7 мкм. Пространство внутри митохондрий заполнено матриксом. Матрикс - это основное вещество митохондрий. В него погружены кольцевая молекула ДНК и рибосомы. Наружная мембрана митохондрий гладкая, она непроницаема для многих веществ. Внутренняя мембрана имеет выросты - кристы, увеличивающие площадь поверхности мембран для протекания химических реакций (рис. 2.32). На поверхности мембраны расположены многочисленные белковые комплексы, составляющие так называемую дыхательную цепь, а также грибовидные ферменты АТФ-синтетазы. В митохондриях протекает аэробный этап дыхания, в ходе которого происходит синтез АТФ.

Пластиды - крупные двумембранные органоиды, характерные только для растительных клеток. Внутреннее пространство пластид заполнено стромой, или матриксом. В строме находится более или менее развитая система мембранных пузырьков - тилакоидов, которые собраны в стопки - граны, а также собственная кольцевая молекула ДНК и рибосомы. Различают четыре основных типа пластид: хлоропласты, хромопласты, лейкопласты и пропластиды.
Хлоропласты - это зеленые пластиды диаметром 3-10 мкм, хорошо различимые под микроскопом (рис. 2.33). Они содержатся только в зеленых частях растений - листьях, молодых стеблях, цветках и плодах. Хлоропласты в основном имеют овальную или эллипсоидную формы, но могут быть также чашевидными, спиралевидными и даже лопастными. Количество хлоропластов в клетке в среднем составляет от 10 до 100 штук.

Однако, например, у некоторых водорослей он может быть один, иметь значительные размеры и сложную форму - тогда его называют хрома- тофором. В других случаях количество хлоропластов может достигать нескольких сотен, при этом их размеры невелики. Окраска хлоропластов обусловлена основным пигментом фотосинтеза - хлорофиллом, хотя в них содержатся и дополнительные пигменты - каротиноиды. Кароти- ноиды становятся заметными только осенью, когда хлорофилл в стареющих листьях разрушается. Основной функцией хлоропластов является фотосинтез. Световые реакции фотосинтеза протекают на мембранах тилакоидов, на которых закреплены молекулы хлорофилла, а темновые реакции - в строме, где содержатся многочисленные ферменты.
Хромопласты. - это желтые, оранжевые и красные пластиды, содержащие пигменты каротиноиды. Форма хромопластов может также существенно варьировать: они бывают трубчатыми, сферическими, кристаллическими и др. Хромопласты придают окраску цветкам и плодам растений, привлекая опылителей и распространителей семян и плодов.
Лейкопласты - это белые или бесцветные пластиды в основном округлой или овальной формы. Они распространены в нефотосинтезирующих частях растений, например в кожице листа, клубнях картофеля и т. д. В них откладываются в запас питательные вещества, чаще всего крахмал, но у некоторых растений это могут быть белки или масло.
Пластиды образуются в растительных клетках из пропластид, которые имеются уже в клетках образовательной ткани и представляют собой небольшие двумембранные тельца. На ранних этапах развития разные виды пластид способны превращаться друг в друга: при попадании на свет лейкопласты клубня картофеля и хромопласты корнеплода моркови зеленеют.
Пластиды и митохондрии называют полуавтономными органоидами клетки, так как они имеют собственные молекулы ДНК и рибосомы, осуществляют синтез белка и делятся независимо от деления клеток. Эти особенности объясняются происхождением от одноклеточных прокариотических организмов. Однако «самостоятельность» митохондрий и пластид является ограниченной, так как их ДНК содержит слишком мало генов для свободного существования, остальная же информация закодирована в хромосомах ядра, что позволяет ему контролировать данные органоиды.
Эндоплазматическая сеть (ЭПС), или эндоплазматический ре тикулум (ЭР) - это одномембранный органоид, представляющий собой сеть мембранных полостей и канальцев, занимающих до 30% содержимого цитоплазмы. Диаметр канальцев ЭПС составляет около 25-30 нм. Различают два вида ЭПС - шероховатую и гладкую. Шероховатая ЭПС несет рибосомы, на ней происходит синтез белков (рис. 2.34).

Гладкая ЭПС лишена рибосом. Ее функция - синтез липидов и углеводов, образование лизосом, а также транспорт, запасание и обезвреживание токсических веществ. Она особенно развита в тех клетках, где происходят интенсивные процессы обмена веществ, например в клетках печени - гепатоцитах - и волокнах скелетных мышц. Вещества, синтезированные в ЭПС, транспортируются в аппарат Гольджи. В ЭПС происходит также сборка мембран клетки, однако их формирование завершается в аппарате Гольджи.
Аппарат Гольджи, или комплекс Гольджи - одномембранный органоид, образованный системой плоских цистерн, канальцев и от- шнуровывающихся от них пузырьков (рис. 2.35).

Структурной единицей аппарата Гольджи является диктиосома - стопка цистерн, наодин полюс которой приходят вещества из ЭПС, а с противоположного полюса, подвергшись определенным превращениям, они упаковываются в пузырьки и направляются в другие части клетки. Диаметр цистерн - порядка 2 мкм, а мелких пузырьков - около 20-30 мкм. Основные функции комплекса Гольджи - синтез некоторых веществ и модификация (изменение) белков, липидов и углеводов, поступающих из ЭПС, окончательное формирование мембран, а также транспорт веществ по клетке, обновление ее структур и образование лизосом. Свое название аппарат Гольджи получил в честь итальянского ученого Камилло Гольджи, впервые обнаружившего данный органоид (1898).
Лизосомы - небольшие одномембранные органоиды до 1 мкм в диаметре, в которых содержатся гидролитические ферменты, участвующие во внутриклеточном пищеварении. Мембраны лизосом слабопроницаемы для этих ферментов, поэтому выполнение лизосомами своих функций происходит очень точно и адресно. Так, они принимают активное участие в процессе фагоцитоза, образуя пищеварительные вакуоли, а в случае голодания или повреждения определенных частей клетки переваривают их, не затрагивая иных. Недавно была открыта роль лизосом в процессах клеточной гибели.
Вакуоль - это полость в цитоплазме растительных и животных клеток, ограниченная мембраной и заполненная жидкостью. В клетках простейших обнаруживаются пищеварительные и сократительные вакуоли. Первые принимают участие в процессе фагоцитоза, так как в них происходит расщепление питательных веществ. Вторые обеспечивают поддержание водно-солевого баланса за счет осморегуляции. У многоклеточных животных в основном встречаются пищеварительные вакуоли.
В растительных клетках вакуоли присутствуют всегда, они окружены специальной мембраной и заполнены клеточным соком. Мембрана, окружающая вакуоль, по химическому составу, строению и выполняемым функциям близка к плазматической мембране. Клеточный сок представляет собой водный раствор различных неорганических и органических веществ, в том числе минеральных солей, органических кислот, углеводов, белков, гликозидов, алкалоидов и др. Вакуоль может занимать до 90 % объема клетки и оттеснять ядро на периферию. Эта часть клетки выполняет запасающую, выделительную, осмотическую, защитную, лизосомную и другие функции, поскольку в ней накапливаются питательные вещества и отходы жизнедеятельности, она обеспечивает поступление воды и поддержание формы и объема клетки, а также содержит ферменты расщепления многих компонентов клетки. К тому же биологически активные вещества вакуолей способны препятствовать поеданию этих растений многими животными. У ряда растений за счет разбухания вакуолей происходит рост клетки растяжением.
Вакуоли имеются также и в клетках некоторых грибов и бактерий, однако у грибов они выполняют только функцию осморегуляции, а у цианобактерий поддерживают плавучесть и участвуют в процессах усвоения азота из воздуха.
Рибосомы - небольшие немембранные органоиды диаметром 15-20 мкм, состоящие из двух субъединиц - большой и малой (рис. 2.36).

Субъединицы рибосом эукариот собираются в ядрышке, а затем транспортируются в цитоплазму. Рибосомы прокариот, митохондрий и пластид меньше по величине, чем рибосомы эукариот. В состав субъединиц рибосом входят рРНК и белки.

Количество рибосом к клетке может достигать нескольких десятков миллионов: в цитоплазме, митохондриях и пластидах они находятся в свободном состоянии, а на шероховатой ЭПС - в связанном. Они принимают участие в синтезе белка, в частности, осуществляют процесс трансляции - биосинтеза полипептидной цепи на молекуле иРНК. На свободных рибосомах синтезируются белки гиалоплазмы, митохондрий, пластид и собственные белки рибосом, тогда как на прикрепленных к шероховатой ЭПС рибосомах осуществляется трансляция белков для выведения из клеток, сборки мембран, образования лизосом и вакуолей.
Рибосомы могут находиться в гиалоплазме поодиночке или собираться в группы при одновременном синтезе на одной иРНК сразу нескольких полипептидных цепей. Такие группы рибосом называются полирибосомами, или полисомами (рис. 2.37).
Микротрубочки - это цилиндрические полые немембранные органоиды, которые пронизывают всю цитоплазму клетки. Их диаметр составляет около 25 нм, толщина стенки - 6-8 нм. Они образованы многочисленными молекулами белка тубулина, которые сначала формируют 13 нитей, напоминающих бусы, а затем собираются в микротрубочку. Микротрубочки образуют цитоплазматическую сеть, которая придает клетке форму и объем, связывают плазматическую мембрану с другими частями клетки, обеспечивают транспорт веществ по клетке, принимают участие в движении клетки и внутриклеточных компонентов, а также в делении генетического материала. Они входят в состав клеточного центра и органоидов движения - жгутиков и ресничек.
Микрофиламенты, или микронити, также являются немембранными органоидами, однако они имеют нитевидную форму и образованы не тубулином, а актином. Они принимают участие в процессах мембранного транспорта, межклеточном узнавании, делении цитоплазмы клетки и в ее движении. В мышечных клетках взаимодействие актиновых микрофиламентов с миозино- выми нитями обеспечивает сокращение.
Микротрубочки и микрофиламенты образуют внутренний скелет клетки - цитоскелет. Он представляет собой сложную сеть волокон, обеспечивающих механическую опору для плазматической мембраны, определяет форму клетки, расположение клеточных органоидов и их перемещение в процессе деления клетки (рис. 2.38).

Клеточный центр - немембранный органоид, располагающийся в животных клетках вблизи ядра; в растительных клетках он отсутствует (рис. 2.39). Его длина составляет около 0,2-0,3 мкм, а диаметр - 0,1-0,15 мкм. Клеточный центр образован двумя центриолями, лежащими во взаимно перпендикулярных плоскостях, и лучистой сферой из микротрубочек. Каждая центриоль образована девятью группами микротрубочек, собранных по три, т. е. триплетами. Клеточный центр принимает участие в процессах сборки микротрубочек, делении наследственного материала клетки, а также в образовании жгутиков и ресничек.

Органоиды движения. Жгутики и реснички представляют собой выросты клетки, покрытые плазмалеммой. Основу этих органоидов составляют девять пар микротрубочек, расположенных по периферии, и две свободные микротрубочки в центре (рис. 2.40). Микротрубочки связаны междусобой различными белками, обеспечивающими их согласованное отклонение от оси - колебание. Колебания энергозависимы, то есть на этот процесс тратится энергия макроэргических связей АТФ. Расщепление АТФ является функцией базальных телец, или кинетосом, расположенных в основании жгутиков и ресничек.
Длина ресничек составляет около 10-15 нм, а жгутиков - 20-50 мкм. За счет строго направленных движений жгутиков и ресничек осуществляется не только движение одноклеточных животных, сперматозоидов и др., но и происходит очистка дыхательных путей, продвижение яйцеклетки по маточным трубам, поскольку все эти части организма человека выстланы реснитчатым эпителием.
Органоиды (органеллы) - это специализированные участки цитоплазмы клетки, имеющие специфическую структуру и выполняющие определенные функции в клетке. Большинство органоидов имеет мембранное строение. Мембраны отсутствуют в структуре рибосом и клеточного центра.
РИБОСОМЫ представляют собой мелкие сферические органоиды, состоящие из двух неравных субъединиц и содержащие примерно равное количество белка и р-РНК. Субъединицы рибосом синтезируются в ядрышках и через поры ядерной мембраны поступают в цитоплазму, где располагаются либо на мембранах эндоплазматической сети, либо свободно. При синтезе белков они могут объединяться на информационной РНК в группы (полисомы) числом от 5 до 70. Рибосомы непосредственно участвуют в сборке белковых молекул. Они содержатся в клетках всех типов.
ЦЕНТРОСОМА, ИЛИ КЛЕТОЧНЫЙ ЦЕНТР - органоид, находящийся вблизи ядра, характерен для большинства животных клеток, имеется у некоторых грибов, водорослей, мхов и папоротников. Это центр организации микротрубочек. Функция центросомы заключается в образовании полюсов деления и формировании микротрубочек веретена деления, с помощью которых происходит растягивание дочерних хромосом в анафазе мейоза и митоза. Хотя центросома играет важнейшую роль в клеточном делении, недавно было показано, что она не является необходимой. У многих живых организмов (животных и ряда простейших) центросома содержит пару центриолей, цилиндрических структур, расположенных под прямым углом друг к другу.
Впервые обнаружена в 1888 г. Теодором Бовери, который назвал её «особым органом клеточного деления». В подавляющем большинстве случаев в клетке в норме присутствует только одна центросома. Аномальное увеличение числа центросом характерно для раковых клеток.
Помимо участия в делении ядра, центросома играет важную роль в формировании жгутиков и ресничек. Центриоли, расположенные в ней, выполняют функцию центров организации для микротрубочек аксонем жгутиков. У организмов, лишенных центриолей (например, у сумчатых и базидиевых грибов, покрытосеменных растений), жгутики не развиваются.
КОМПЛЕКС (АППАРАТ) ГОЛЬДЖИ - сложная сеть, расположенная вокруг ядра (сетчатый комплекс). В клетках протистов и растений он представлен отдельными тельцами серповидной или палочковидной формы – диктиосомами, каналами, цистернами, которые окружены мембранами. В них происходит сортировка и упаковка поступивших макромолекул. От них отпочковываются пузырьки с веществами, необходимыми клетке. Комплекс Гольджи соединен с каналами эндоплазматической сети. Основные его функции: 1) концентрация, обезвоживание и уплотнение синтезированных в клетке белков, жиров, полисахаридов и веществ, поступивших извне, подготовка их к использованию или выведению из клетки; 2) образование лизосом и сборка сложных комплексов органических веществ, например гликопротеинов.
ЛИЗОСОМЫ - шаровидные мелкие тельца (пузырьки), покрытые элементарной мембраной и содержащие около 40 гидролитических ферментов, способных в кислой среде (рН 4,5-5,0) расщеплять белки, нуклеиновые кислоты, жиры и углеводы. Лизосомы могут также лизировать стареющие органоиды. Образование лизосом происходит в комплексе Гольджи. Продукты лизиса через мембрану лизосом поступают в цитоплазму и включаются в дальнейший обмен веществ.
СФЕРОСОМЫ - мелкие тельца, первоначально окруженные биологической мембраной и содержащие специфические ферменты. Функция сферосом состоит в накоплении жира. Зрелая сферосома обычно представляет собой каплю жира, окруженную биологической мембраной или белковой оболочкой.
Мелкие сферические или эллипсоидальные органоиды, окруженные одной мембраной, получили название микротелец . Наиболее известные из них - глиоксисомы и пероксисомы.
ГЛИОКСИСОМЫ содержат ферменты, необходимые для превращения жиров в углеводы, что происходит во время прорастания семян. В них осуществляется цикл глиоксиловой кислоты.
ПЕРОКСИСОМЫ обнаружены в большинстве типов клеток. Функции пероксисом зависят от типа клетки. В ряде случаев они имеют непосредственное отношение к фотодыханию, играя важную роль в метаболизме гликолевой кислоты.
ПАРАМУРАЛЬНЫЕ ТЕЛЬЦА - особые тельца, первоначально возникающие в виде впячиваний в плазмалемме. Такие впячивания позднее могут отделяться от плазмалеммы и внедряться в цитоплазму.
ПЛАЗМИДЫ представляют собой кольцевые двуцепочечные молекулы ДНК, существующие в большинстве изученных клеток в автономном, не связанном с хромосомами состоянии. Они являются внехромосомными факторами наследственности и интенсивно используются в генной инженерии в качестве молекулярных переносчиков чужеродной ДНК. Наиболее изучены бактериальные плазмиды.
ОРГАНОИДЫ ДВИЖЕНИЯ КЛЕТОК (у животных) представлены жгутиками и ресничками. Это выросты цитоплазмы, покрытые элементарной мембраной, под которой находится 20 микротрубочек, образующих 9 пар по периферии и 2 одиночные в центре. У основания ресничек и жгутиков расположены базальные тельца, образующие микротрубочки этих органоидов. Длина жгутиков достигает 100 мкм. Короткие (10-20 мкм) многочисленные жгутики называются ресничками. Реснички и жгутики служат для передвижения организмов (бактерии, протесты, ресничные черви), половых клеток (сперматозоидов) либо для перемещения частиц или жидкостей (реснички мерцательного эпителия дыхательных путей, яйцеводов и др.).
МИТОХОНДРИИ - это палочковидные, нитевидные или шаровидные органеллы. Оболочка митохондрий состоит из двух мембран - наружной гладкой, и внутренней, образующейвыросты - кристы, кармановидные мешочки, которые вдаются во внутреннее гомогенное содержимое митохондрии - матрикс. Совокупность митохондрий в клетке называют хондриомом .
Наружная мембрана проницаема для неорганических ионов и относительно крупных молекул, в частности аминокислот, сахарозы и т. д., регулирует поступление веществ в митохондрию и их выведение.
В матриксе встречаются рибосомы, митохондриальная ДНК, промежуточные продукты обмена, а также многочисленные ферменты, которые локализованы на внутренней мембране, за счет этого резко увеличивается поверхность митохондрии. Митохондрии являются дыхательными центрами клетки, присутствуют во всех клетках с аэробным дыханием.
Основная функция митохондрии - образование энергии. Большая часть энергии сразу же расходуется на синтез АТФ из АДФ, часть непосредственно используется на активный транспорт через мембрану или на выработку тепла. Молекулы богатой энергией АТФ выходят из митохондрии и используются для поддержания процессов жизнедеятельности клетки - поглощения, выделения, различных синтезов, деления и т. д. При этом АТФ превращается в АДФ, которая опять поступает в митохондрии.
Источником энергии являются процессы окисления различных веществ (главным образом сахаров). Окисление, осуществляющееся в растительной клетке в процессе дыхания, сопровождается выделением большого количества энергии, которая консервируется в митохондриях путем образования АТФ. Присоединение остатка фосфорной кислоты к АДФ при синтезе АТФ в митохондриях носит название окислительного фосфорилирования.
Митохондрии могут делиться пополам (перешнуровываться) либо отпочковываться. В клетке митохондрии развиваются под контролем ядра.
ПЛАСТИДЫ - органоиды, содержащиеся только в растительных клетках. Они подразделяются на три группы - хлоропласты (зеленые), хромопласты (чаще желтые или оранжевые) и лейкопласты (бесцветные). Предшественниками пластид являются протопластиды (этиопласты) – бесцветные образования в делящихся клетках. Пластиды имеют сходное строение и при определенных условиях могут переходить из одного вида в другой. Так, при хранении картофеля и моркови на свету лейкопласты и хромопласты превращаются в хлоропласты (овощи зеленеют). Совокупность всех пластид в клетке называют пластидомом .
Хлоропласты по форме напоминают двояковыпуклую линзу и содержат зеленый пигмент хлорофилл. Существует несколько модификаций хлорофиллов - а, b, c, d. Хлоропласты имеются в листьях, молодых побегах, незрелых плодах. Стенка хлоропласта образована двумя мембранами , внутри находится бесструктурное содержимое - строма. Строма пронизана системой параллельно расположенных элементарных мембран, являющихся продолжением внутренней мембраны. Их называют тилакоидами . В некоторых местах мембраны тилакоидов плотно прилегают друг к другу, образуя стопки - граны. На тилактоидах гран располагаются молекулы хлорофилла, улавливающие солнечный свет, и ферменты, синтезирующие АТФ. В строме локализованы ферменты фиксации СО 2 и синтеза органических соединений с использованием энергии АТФ. Таким образом, в гранах протекает световая фаза фотосинтеза, а в строме - темновая. В строме хлоропластов имеется автономная система синтеза белков (ДНК, РНК и рибосомы). Основные функции хлоропластов - фотосинтез и синтез специфических белков. У водорослей хлоропласт часто один, крупный, специфический и называется хроматофором .
Лейкопласты - бесцветные пластиды, содержащиеся чаще в неокрашенных частях растений - стеблях, корнях, луковицах и т. п. Форма их может быть различной и непостоянна, внутренние мембраны развиты слабо. В лейкопластах могут синтезироваться и накапливаться белки, жиры и полисахариды (крахмал). Лейкопласты, накапливающие крахмал, называют амилопластами, накапливающие белки – протеопластами , жирные масла - олеопластами .
Хромопласты - пластиды, содержащие растительные пигменты (кроме зеленого), придающие окраску цветкам, плодам, стеблям и другим частям растений благодаря накоплению в них каротиноидов. Хромопласты – конечный этап в развитии пластид. Они мельче хлоропластов, имеют нелинзовидную форму, внутренняя мембранная система у них обычно отсутствует. Наиболее часто хлоропласты превращаются в хромопласты при осеннем пожелтении листьев или созревании плодов. Процесс превращения других пластид в хромопласты необратим.
ЯДРО РАСТИТЕЛЬНОЙ КЛЕТКИ является обязательным компонентом всех растительных эукариотических клеток. Некоторые клетки имеют два и более ядер (грибы и др.). Форма и размеры ядра зависят от формы и величины клетки и выполняемой ею функции. В округлых и многоугольных клетках оно обычно шаровидное, в вытянутых - палочковидное или овальное.
По химическому составу ядро отличается от остальных компонентов клетки высоким содержанием ДНК (15-30 %) и РНК (12 %); 99 % ДНК клетки сосредоточено в ядре, где она вместе с белками образует комплексы - дезоксирибонуклеопротеины (ДНП).
Ядро выполняет две главные функции:
♦ хранение и воспроизведение наследственной информации;
♦ регуляция процессов обмена веществ, протекающих в клетке.
В процессе деления клеток структуры ядра претерпевают значительные изменения.
В интерфазном ядре
различают ядерную оболочку, ядерный сок, хроматин и ядрышки.
Ядерная оболочка (кариолемма) представлена двумя биологическими мембранами, между которыми находится перинуклеарное пространство. Наружная ядерная мембрана непосредственно соединена с мембранами каналов эндоплазматической сети. На наружной мембране располагаются рибосомы, внутренняя мембрана гладкая. Ядерная оболочка пронизана многочисленными порами, через которые происходит обмен веществ между ядром и цитоплазмой. Основная функция ядерной оболочки - регуляция обмена веществ. Кроме того, она выполняет защитную функцию.
Ядерный сок (кариоплазма) - это однородная масса, заполняющая пространство между структурами ядра (хроматином и ядрышками). В его состав входят вода, белки (ферменты), нуклеотиды, аминокислоты и различные виды РНК (и-РНК, т-РНК, р-РНК). Ядерный сок осуществляет взаимосвязь ядерных структур и обмен с цитоплазмой клетки.
Хроматин представляет собой дезоксирибонуклеопротеин (ДНП), выявляемый под световым микроскопом в виде тонких нитей и гранул. Так выглядят деспирализованные хромосомы в интерфазе. В процессе митоза хроматин путем спирализации образует хорошо видимые (особенно в метафазе) интенсивно окрашивающиеся структуры - хромосомы. Основная функция хромосом состоит в хранении, воспроизведении и передаче генетической информации в клетке.
Метафазная хромосома состоит из двух продольных нитей ДНП - хроматид, соединенных друг с другом в области первичной перетяжки - центромеры, к которой прикрепляются нити веретена деления. Центромера делит тело хромосомы на два плеча. В зависимости от расположения первичной перетяжки различают следующие типы хромосом: метацентрические (равноплечие,), в которых центромера расположена посередине, а плечи примерно равной длины; субметацентрические (неравноплечие), когда центромера смещена от середины хромосомы, а плечи неравной длины; акроцентрические (палочковидные,), когда центромера смещена к концу хромосомы и одно плечо очень короткое. В некоторых хромосомах могут быть вторичные перетяжки, отделяющие от тела хромосомы участок, называемый спутником .
Ядрышки обычно имеют шаровидную форму, не окружены мембраной и находятся в контакте с ядерным соком. Они содержат белки и р-РНК в равном соотношении. Ядрышки - непостоянные образования, они растворяются в начале деления клетки и восстанавливаются после его окончания. Их образование связано со вторичными перетяжками (ядрышковыми организаторами) спутничных хромосом. В области вторичных перетяжек локализованы гены, кодирующие синтез рибосомальной РНК и белков. В ядрышках происходит формирование рибосом, которые затем выходят в цитоплазму через поры в ядерной оболочке.
РАЗЛИЧИЯ РАСТИТЕЛЬНОЙ И ЖИВОТНОЙ КЛЕТКИ:
♦ клетки животных не имеют клеточной стенки (покрыты только элементарной мембраной), у клеток растений есть клеточная стенка (поверх мембраны имеется оболочка: у растений ее основой является полисахарид целлюлоза, у грибов стенка состоит главным образом из азотсодержащего полисахарида хитина). Симпластический обмен веществ осуществляется в растительных клетках осуществляется через плазмодесмы.
♦ животная клетка - гетеротроф, она не содержит пластид, растительная - автотроф, имеет пластиды;
♦ в животной клетке имеются центриоли, в растительной - нет;
♦ в животной клетке нет центральной вакуоли, в растительной она имеется и содержит клеточный сок;
запасное питательное вещество животной клетки и у большинства грибов - гликоген,
у растительной – полисахарид крахмал.
ДЕЛЕНИЕ КЛЕТОК. У многоклеточных организмов рост и развитие происходят в результате роста и деления клеток, составляющих его тело. Существуют 4 способа деления клетки: амитоз, эндомитоз, митоз и мейоз.
АМИТОЗ, или прямое деление, - способ, при котором происходит вначале разделение ядрышка, затем простое деление ядра на два перетяжкой в виде цифры 8, вслед за этим наступает полное деление протопласта и всей клетки на две. При этом ядерное вещество не всегда равномерно распределяется между дочерними клетками. Амитоз встречается в клетках стареющих тканей либо больных, поэтому деление носит патологический характер. Открыт Николаем Ивановичем Железновым в 1840 году.
ЭНДОМИТОЗ представляет собой внутриклеточное деление. Происходит редупликация хромосом в клетке, но хромосомы не расходятся по полюсам. Эндомитоз часто является причиной полиплоидии.
МИТОЗ , или кариокинез, - широко распространенный, универсальный способ деления. Этим способом делятся вегетативные клетки (соматические) всех растений, животных и человека. Митотическое деление - сложный процесс, благодаря которому клеточный материал распределяется между дочерними клетками поровну. Открыт в 1874 году Иваном Дорофеевичем Чистяковым.
Митоз - одна из частей клеточного цикла, но так как он достаточно сложен, в его составе были выделены четыре фазы: профаза, метафаза, анафаза и телофаза. Удвоение хромосом происходит еще в ходе интерфазы. В результате этого, в митоз хромосомы вступают уже удвоенными, напоминающими букву X (идентичные копии материнской хромосомы соединены друг с другом в области центромеры). Продолжительность митоза составляет 0,5-3 ч.
В профазе начинает увеличиваться объем ядра, хромосомы становятся видимыми вследствие спирализации хроматина. К концу профазы заметно, что каждая хромосома состоит из двух хроматид, соединенных в области центромеры. Постепенно исчезает ядрышко, разрушается ядерная оболочка, образуется веретено деления.
Метафаза характеризуется максимальной спирализацией хромосом. Они упорядочение располагаются на экваторе клетки, образуя метафазную пластинку. При этом отчетливо видно, что каждая хромосома состоит из двух хроматид (2n2хр), поэтому подсчет и изучение хромосом проводят именно в данный период.
В анафазе соединение в районе центромеры разрушается, хромосомы делятся и расходятся к полюсам деления.
В телофазу собравшиеся у полюсов хромосомы деспирализуются и становятся плохо видимыми. Вокруг них из мембранных структур цитоплазмы образуется ядерная оболочка. Восстанавливаются ядрышки. Одновременно идет деление цитоплазмы в животных клетках - путем перетяжки, а в растительных - путем построения мембраны, начиная с середи ны клетки (цитокинез). Образовавшиеся дочерние клетки имеют диплоидный набор хромосом, каждая из которых состоит из одной хроматиды (2n1хр).
Биологическое значение митоза заключается в точном распределении хромосом и содержащейся в них генетической информации между дочерними клетками, что обеспечивает постоянство кариотипа и генетическую преемственность в многочисленных клеточных поколениях. Митоз обусловливает важнейшие явления жизнедеятельности: рост, развитие и восстановление тканей и органов организма.
МЕЙОЗ (редукционное деление). Открыт в 1885 году Владимиром Ивановичем Беляевым. Мейозу подвержены половые клетки (гаметы). Весь процесс слагается из двух делений ядра, быстро следующих друг за другом. Наиболее сложно первое деление, при котором происходит редукция хромосом. Второе деление протекает как типичное митотическое. В результате мейоза образуются 4 гаплоидные клетки, представляющие в одних случаях споры (у большинства низших и у всех высших архегониальных растений), а в других - гаметы.
Профаза мейоза I длительна и распадается на 5 стадий – лептонему, зигонему, пахинему диплонему, диакинез. Происходит постепенная спирализация хроматина, образуются видимые хромосомы. Гомологичные хромосомы попарно сближаются сначала в области центромер, а затем по всей длине, образуя одну общую структуру, состоящую из двух хромосом и четырех хроматид. Их называют бивалентами или тетрадами (bi - два, tetra - четыре). Тесное соприкосновение двух гомологичных хромосом называется конъюгацией. В процессе конъюгации между некоторыми хроматидами гомологичных хромосом может происходить обмен участками - кроссинговер, приводящий к перекомбинации генетического материала. К концу профазы растворяются ядерная оболочка и ядрышки, формируется ахроматиновое веретено деления. Проконъюгировавшие хромосомы сначала разделяются в области центромер, оставаясь соединенными в области плеч, и образуют перекресты (хиазмы). Расхождение хроматид постепенно увеличивается, и перекресты смещаются к их концам. Содержание генетического материала в этот период - 2n2хр.
В метафазе мейоза I гомологичные хромосомы располагаются попарно в экваториальной плоскости клетки. В этот момент спирализация их достигает максимума. Содержание генетического материала не изменяется (2n2хр).
В анафазе мейоза I гомологичные хромосомы, состоящие из двух хроматид, расходятся к противоположным полюсам клетки. Следовательно, из каждой пары гомологичных хромосом в дочернюю клетку попадает только одна - число хромосом уменьшается вдвое (происходит редукция). Содержание генетического материала становится 1n2хр у каждого полюса.
В телофазе происходит формирование ядер и разделение цитоплазмы - образуются две дочерние клетки. Каждая клетка содержит гаплоидный набор хромосом, состоящих из двух хроматид (1n2хр).
Интеркинез – переходная фаза между делениями.
Мейоз II протекает по типу митоза. В метафазе хромосомы располагаются в экваториальной плоскости клетки. Изменений генетического материала не происходит (1n2хр). В анафазе мейоза II хроматиды каждой хромосомы отходят к противоположным полюсам клетки и содержание генетического материала у каждого полюса становится 1n1хр. В телофазе образуются 4 гаплоидные клетки (1n1хр).
Редукционное деление имеет важное биологическое значение . 1) Благодаря редукции хромосом сохраняются виды, так как гаметы с гаплоидным числом хромосом после слияния восстанавливают первоначальное число хромосом, характерное для данного вида. 2) Обеспечивается возможность рекомбинации хромосом и генов при половом процессе. Это обеспечивает появление разнообразного и разнокачественного потомства при половом размножении организмов. 3) Благодаря мейозу происходит чередование ядерных фаз - диплоидной и гаплоидной, что, в свою очередь, обусловливает чередование бесполого (спорофита) и полового (гаметофита) поколения в цикле их развития. Чередование поколений играет решающую роль в сохранении видов, образованных в результате эволюции.

Митоз . I-III - профаза; IV - метафаза;
V-VI - анафаза; VII-VIII - телофаза.

Мейоз
. Профаза I (1-5), 6 - метафаза I;
7 - анафаза I; 8 - телофаза I; 9 - интеркинез;
10 - метафаза II; 11 - анафаза II; 12 - телофаза II.
Одна из двух гомологичных хромосом заштрихована, другая - белая. Обмен белыми
и заштрихованными участками хромосом - результат кроссинговера.
Маленькие белые кружки - центромеры, большой круг - контур ядра.
В метафазе и анафазе обоих делений ядерная мембрана исчезает. В телофазе возникает снова. В метафазе и анафазе обоих делений стрелками показано направление растягивания и движения хромосом с помощью нитей веретена.
Кристаллы и скопления минеральных солей в клетках:
1 - цистолит в клетке эпидермы листа инжира, 2 - рафиды в клетках листа традесканции, 3 - друзы в клетках палисадной ткани листа инжира, 4 - друзы и одиночные кристаллы в клетках черешка бегонии, 5 - одиночные кристаллы в клетках эпидермы чешуи луковицы лука, 6 - скопление мелких кристаллов («кристаллический песок») в клетках мезофилла листа красавки (белладонны)
Целлюлоза (клетчатка) . Она, как и крахмал, является полимером глюкозы, однако из-за различий в строении молекулярной цепочки целлюлоза не расщепляется в кишечнике человека.
Пектин - природный полимер Д - галактуроновой кислоты
Гемицеллюлоза – полисахарид клеточной оболочки, состоящий из полимеров глюкозы и гексозы. Г. отличается от целлюлозы лучшей р-римостью в р-рах щелочей и способностью легко гидролизоваться кипящими разб. минеральными к-тами.
Молекула лигнина состоит из продуктов полимеризации ароматических спиртов.
Все простейшие - одноклеточные или многоклеточные, не имеющие высокоорганизованных тканей.
Мононуклеотид аденозинтрифосфорная кислота, аденозинтрифосфат, состоящий из азотистого основания аденина, пятиуглеродного моносахарида рибозы и трех остатков фосфорной кислоты, которые соединены между собой высокоэнергетическими связями
Центриоли принимают участие в формировании цитоплазматических микротрубочек во время деления клетки и в регуляции образования митотического веретена. В клетках растений центриолей нет, и митотическое веретено образуется там иным способом.
Архегониальные растения (Archegoniatae), растения, имеющие женский половой орган в форме архегония. А. р. впервые были выделены в отдельный тип в 1876 русским ботаником И.Н. Горожанкиным, который включал сюда голосеменные, мохообразные и папоротникообразные, в отличие от покрытосеменных, у которых нет архегония, а имеется сложный женский орган - пестик. Большинство ботаников выделяют эти группы в три самостоятельных типа: мохообразные, папоротникообразные и голосеменные.
Органоиды клетки, они же органеллы, представляют собой специализированные структуры собственно клетки, отвечающие за различные важные и жизненно необходимые функции. Почему же все-таки «органоиды»? Просто тут эти компоненты клетки сопоставляются с органами многоклеточного организма.
Какие органоиды входят в состав клетки
Также порой под органоидами понимается исключительно лишь постоянные структуры клетки, которые находятся в ее . По этой же причине ядро клетки и ее ядрышко не называют органоидами, равно как и не являются органоидами , реснички и жгутики. А вот к органоидам, входящим в состав клетки относятся: , комплекс , эндоплазматическая сеть, рибосомы, микротрубочки, микрофиламенты, лизосомы. По сути это и есть основные органоиды клетки.
Если речь идет о животных клетках, то в число их органоидов также входят центриоли и микрофибриллы. А вот в число органоидов растительной клетки еще входят только свойственные растениям пластиды. В целом состав органоидов в клетках может существенно отличатся в зависимости от вида самой клетки.

Рисунок строения клетки, включая ее органоиды.
Двумембраные органоиды клетки
Также в биологии существует такое явление как двумембраные органоиды клетки, к ним относятся митохондрии и пластиды. Ниже мы опишем свойственные им функции, впрочем, как всех других основных органоидов.
Функции органоидов клетки
А теперь коротко опишем основные функции органоидов животной клетки. Итак:
- Плазматическая мембрана – тонкая пленка вокруг клетки состоящая из липидов и белков. Очень важный органоид, который обеспечивает транспортировку в клетку воды, минеральных и органических веществ, удаляет вредные продукты жизнедеятельности и защищает клетку.
- Цитоплазма – внутренняя полужидкая среда клетки. Обеспечивает связь между ядром и органоидами.
- Эндоплазматическая сеть – она же сеть каналов в цитоплазме. Принимает активное участие в синтезе белков, углеводов и липидов, занимается транспортировкой полезных веществ.
- Митохондрии – органоиды, в которых окисляются органические вещества и синтезируются молекулы АТФ с участием ферментов. По сути митохондрии это органоид клетки, синтезирующий энергию.
- Пластиды (хлоропласты, лейкопласты, хромопласты) – как мы упоминали выше, встречаются исключительно у растительных клеток, в целом их наличие является главной особенностью растительного организма. Играют очень важную функцию, например, хлоропласты, содержащие зеленый пигмент хлорофилл, у растения отвечают за явление .
- Комплекс Гольджи — система полостей, отграниченных от цитоплазмы мембраной. Осуществляют синтез жиров и углеводов на мембране.
- Лизосомы - тельца, отделенные от цитоплазмы мембраной. Имеющиеся в них особые ферменты ускоряют реакцию расщепления сложных молекул. Также лизосома является органоидом, обеспечивающим сборку белка в клетках.
- - полости в цитоплазме, заполненные клеточным соком, место накопления запасных питательных веществ; они регулируют содержание воды в клетке.
В целом все органоиды являются важными, ведь они регулируют жизнедеятельность клетки.
Основные органоиды клетки, видео
И в завершение тематическое видео про органоиды клетки.
Органеллами (органоидами) клетки называют постоянные части клетки, имеющие определённое строение и выполняющие специфические функции. Различают мембранные и немембранные органеллы. К мембранным органеллам относят цитоплазматическую сеть (эндоплазматический ретикулум), пластинчатый комплекс (аппарат Гольджи), митохондрии, лизосомы, пероксисомы. Немембранные органеллы представлены рибосомами (полирибосомами), клеточным центром и элементами цитоскелета: микротрубочками и фибриллярными структурами.
Рис. 8. Схема ультрамикроскопического строения клетки:
1 – гранулярная эндоплазматическая сеть, на мембранах которой расположены прикреплённые рибосомы; 2 – агранулярная эндоплазматическая сеть; 3 – комплекс Гольджи; 4 – митохондрия; 5 – формирующаяся фагосома; 6 – первичная лизосома (гранула накопления); 7 – фаголизосома; 8 – эндоцитозные пузырьки; 9 – вторичная лизосома; 10 – остаточное тельце; 11 – пероксисома; 12 – микротрубочки; 13 - микрофиламенты; 14 – центриоли; 15 – свободные рибосомы; 16 – транспортные пузырьки; 17 – экзоцитозный пузырёк; 18 – жировые включения (липидная капля); 19 - включения гликогена; 20 – кариолемма (ядерная оболочка); 21 – ядерные поры; 22 – ядрышко; 23 – гетерохроматин; 24 – эухроматин; 25 – базальное тельце реснички; 26 - ресничка; 27 – специальный межклеточный контакт (десмосома); 28 – щелевой межклеточный контакт
2.5.2.1. Мембранные органоиды (органеллы)
Эндоплазматическая сеть (эндоплазматический ретикулум, цитоплазматическая сеть) - совокупность сообщающихся между собой канальцев, вакуолей и «цистерн», стенка которых образована элементарными биологическими мембранами. Открыта К.Р. Портером в 1945 году. Открытие и описание эндоплазматической сети (ЭПС) обязано внедрению в практику цитологических исследований электронного микроскопа. Мембраны, образующие ЭПС, отличаются от плазмолеммы клетки меньшей толщиной (5-7 нм) и большей концентрацией белков, в первую очередь обладающих ферментативной активностью. Различают две разновидности ЭПС (рис. 8): шероховатую (гранулярную) и гладкую (агранулярную). Шероховатая ЭПС представлена уплощенными цистернами, на поверхности которых расположены рибосомы и полисомы. Мембраны гранулярной ЭПС содержат белки, способствующие связыванию рибосом и уплощению цистерн. Особенно хорошо развита шероховатая ЭПС в клетках, специализирующихся на белковом синтезе. Гладкую ЭПС формируют переплетающиеся канальцы, трубочки и небольшие пузырьки. Каналы и цистерны ЭПС этих двух разновидностей не разграничены: мембраны одного типа переходят в мембраны другого типа, формируя в области перехода так называемую переходную (транзиторную) ЭПС.
Основными функциями гранулярной ЭПС являются:
1) синтез на прикреплённых рибосомах белков (секретируемых белков, белков клеточных мембран и специфических белков содержимого мембранных органоидов); 2) гидроксилирование, сульфатирование, фосфорилирование и гликозилирование белков; 3) транспорт веществ в пределах цитоплазмы; 4) накопление как синтезируемых, так и транспортируемых веществ; 5) регуляция биохимических реакций, связанная с упорядоченностью локализации в структурах ЭПС веществ, вступающих в реакции, а также их катализаторов - ферментов.
Гладкая ЭПС отличается отсутствием на мембранах белков (рибофоринов), связывающих субъединицы рибосом. Предполагается, что гладкая ЭПС образуется в результате формирования выростов шероховатой ЭПС, мембрана которых утрачивает рибосомы.
Функциями гладкой ЭПС являются: 1) синтез липидов, включая мембранные липиды; 2) синтез углеводов (гликогена и др.); 3) синтез холестерина; 4) обезвреживание токсических веществ эндогенного и экзогенного происхождения; 5) накопление ионов Са 2+ ; 6) восстановление кариолеммы в телофазе митоза; 7) транспорт веществ; 8) накопление веществ.
Как правило, гладкая ЭПС развита в клетках слабее, чем шероховатая ЭПС, однако в клетках, вырабатывающих стероиды, триглицериды и холестерин, а также в клетках печени, осуществляющих детоксикацию различных веществ, она развита значительно лучше.

Рис. 9. Комплекс Гольджи:
1 – стопка уплощённых цистерн; 2 – пузырьки; 3 – секреторные пузырьки (вакуоли)
Переходная (транзиторная) ЭПС - это участок перехода гранулярной ЭПС в агранулярную ЭПС, который располагается у формирующейся поверхности комплекса Гольджи. Трубочки и канальцы переходной ЭПС распадаются на фрагменты, из которых образуются пузырьки, транспортирующие материал из ЭПС в комплекс Гольджи.
Пластинчатый комплекс (комплекс Гольджи, аппарат Гольджи) - органоид клетки, участвующий в окончательном формировании продуктов её жизнедеятельности (секретов, коллагена, гликогена, липидов и других продуктов), а также в синтезе гликопротеидов. Органоид назван по имени описавшего его в 1898 году итальянского гистолога К. Гольджи. Образован тремя составляющими (рис. 9): 1) стопкой уплощённых цистерн (мешочков); 2) пузырьками; 3) секреторными пузырьками (вакуолями). Зона скопления этих элементов получила название диктиосомы. Таких зон в клетке может быть несколько (иногда несколько десятков и даже сотен). Комплекс Гольджи располагается около ядра клетки, часто вблизи центриолей, реже рассеян по всей цитоплазме. В секреторных клетках он располагается в апикальной части клетки, через которую осуществляется выделение секрета путём экзоцитоза. От 3-х до 30-ти цистерн в виде изогнутых дисков диаметром 0,5-5 мкм образуют стопку. Смежные цистерны разделены пространствами в 15-30 нм. Отдельные группы цистерн в пределах диктиосомы отличаются особым составом ферментов, определяющих характер биохимических реакций, в частности процессинга белка и др.
Второй составляющий элемент диктиосомы - пузырьки представляют собой сферические образования диаметром 40-80 нм, умеренно плотное содержимое которых окружено мембраной. Пузырьки формируются путём отщепления от цистерн.
Третий элемент диктиосомы - секреторные пузырьки (вакуоли) представляют собой относительно крупные (0,1-1,0 мкм) сферические мембранные образования, содержащие секрет умеренной плотности, претерпевающий конденсацию и уплотнение (вакуоли конденсации).
Комплекс Гольджи отчётливо поляризован по вертикали. В нём выделяют две поверхности (два полюса):
1) цис-поверхность, или незрелую поверхность, которая имеет выпуклую форму, обращена к эндоплазматической сети (ядру) и связана с отделяющимися от неё мелкими транспортными пузырьками;
2) транс-поверхность, или поверхность, обращённую к плазмолемме вогнутой формы (рис. 8), со стороны которой от цистерн комплекса Гольджи отделяются вакуоли (секреторные гранулы).
Основными функциями комплекса Гольджи являются: 1) синтез гликопротеинов и полисахаридов; 2) модификация первичного секрета, его конденсация и упаковка в мембранные пузырьки (формирование секреторных гранул); 3) процессинг молекул (фосфорилирование, сульфатирование, ацилирование и т.п.); 4) накопление секретируемых клеткой веществ; 5) образование лизосом; 6) сортировка синтезированных клеткой белков у транс-поверхности перед их окончательным транспортом (производится посредством рецепторных белков, распознающих сигнальные участки макромолекул и направляющих их в различные пузырьки); 7) транспорт веществ: из транспортных пузырьков вещества проникают в стопку цистерн комплекса Гольджи с цис-поверхности, а выходят из неё в виде вакуолей с транс-поверхности. Механизм транспорта объясняют две модели: а) модель перемещения пузырьков, отпочковывающихся от предшествующей цистерны и сливающихся с последующей цистерной последовательно в направлении от цис-поверхности к транс-поверхности; б) модель перемещения цистерн, основанная на представлении о непрерывном новообразовании цистерн за счёт слияния пузырьков на цис-поверхности и последующем распаде на вакуоли цистерн, смещающихся к транс-поверхности.
Указанные выше основные функции позволяют констатировать, что пластинчатый комплекс - важнейший органоид клетки эукариот, обеспечивающий организацию и интеграцию внутриклеточного метаболизма. В этом органоиде протекают заключительные этапы формирования, созревания, сортировки и упаковки всех секретируемых клеткой продуктов, ферментов лизосом, а также белков и гликопротеинов поверхностного аппарата клетки и др. веществ.
Органоиды внутриклеточного переваривания. Лизосомы - это мелкие ограниченные элементарной мембраной пузырьки, содержащие гидролитические ферменты. Мембрана лизосом толщиной около 6 нм осуществляет пассивную компартментализацию, временно отделяя гидролитические ферменты (более 30 разновидностей) от гиалоплазмы. В неповреждённом состоянии мембрана устойчива к действию гидролитических ферментов и препятствует их утечке в гиалоплазму. В стабилизации мембраны важная роль принадлежит кортикостероидным гормонам. Повреждение мембран лизосом ведёт к самоперевариванию клетки гидролитическими ферментами.
Мембрана лизосом содержит АТФ-зависимый протонный насос, обеспечивающий закисление среды внутри лизосом. Последняя способствует активизации ферментов лизосом - кислых гидролаз. Наряду с этим мембрана лизосом содержит рецепторы, обусловливающие связывание лизосом с транспортными пузырьками и фагосомами. Мембрана обеспечивает также диффузию веществ из лизосом в гиалоплазму. Связывание части молекул гидролаз с мембраной лизосом ведёт к их инактивации.
Выделяют несколько разновидностей лизосом: первичные лизосомы (гидролазные пузырьки), вторичные лизосомы (фаголизосомы, или пищеварительные вакуоли), эндосомы, фагосомы, аутофаголизосомы, остаточные тельца (рис. 8).
Эндосомами называют мембранные пузырьки, переносящие макромолекулы от поверхности клетки в лизосомы путём эндоцитоза. В процессе переноса содержимое эндосом может не изменяться или претерпевать частичное расщепление. В последнем случае в эндосомы проникают гидролазы или эндосомы непосредственно сливаются с гидролазными пузырьками, вследствие чего среда постепенно закисляется. Эндосомы разделяют на две группы: ранние (периферические) и поздние (перинуклеарные) эндосомы.
Ранние (периферические) эндосомы формируются на ранних этапах эндоцитоза после отделения пузырьков с захваченным содержимым от плазмолеммы. Они располагаются в периферических слоях цитоплазмы и характеризуются нейтральной или слабощелочной средой. В них происходит отщепление лигандов от рецепторов, сортировка лигандов и, возможно, возвращение рецепторов в специальных пузырьках в плазмолемму. Наряду с этим в ранних эндосомах может происходить расщепление ком-

Рис. 10 (А). Схема образования лизосом и их участия во внутриклеточном пищеварении. (Б) Электронная микрофотография среза вторичных лизосом (обозначены стрелками):
1 – образование из гранулярной эндоплазматической сети мелких пузырьков с ферментами; 2 – перенос ферментов в аппарат Гольджи; 3 – образование первичных лизосом; 4 – выделение и использование (5) гидролаз при внеклеточном ращеплении; 6 - фагосомы; 7 – слияние первичных лизосом с фагосомами; 8, 9 – образование вторичных лизосом (фаголизосом); 10 – экскреция остаточных телец; 11 – слияние первичных лизосом с разрушающимися структурами клетки; 12 – аутофаголизосома
плексов «рецептор-гормон», «антиген-антитело», ограниченное расщепление антигенов, инактивация отдельных молекул. В условиях закисления (рН=6,0) среды в ранних эндосомах может происходить частичное расщепление макромолекул. Постепенно, перемещаясь вглубь цитоплазмы, ранние эндосомы превращаются в поздние (перинуклеарные) эндосомы, располагающиеся в глубоких слоях цитоплазмы, окружающих ядро. Они достигают 0,6-0,8 мкм в диаметре и отличаются от ранних эндосом более кислым (рН=5,5) содержимым и более высоким уровнем ферментативного переваривания содержимого.
Фагосомы (гетерофагосомы) - мембранные пузырьки, которые содержат захваченный клеткой извне материал, подлежащий внутриклеточному перевариванию.
Первичные лизосомы (гидролазные пузырьки) - пузырьки диаметром 0,2-0,5 мкм, содержащие неактивные ферменты (рис.10). Их перемещение в цитоплазме контролируется микротрубочками. Гидролазные пузырьки осуществляют транспорт гидролитических ферментов из пластинчатого комплекса к органоидам эндоцитозного пути (фагосомам, эндосомам и т.п.).
Вторичные лизосомы (фаголизосомы, пищеварительные вакуоли) - пузырьки, в которых активно осуществляется внутриклеточное переваривание посредством гидролаз при рН≤5. Их диаметр достигает 0,5-2 мкм. Вторичные лизосомы (фаголизосомы и аутофаголизосомы) формируются путём слияния фагосомы с эндосомой или первичной лизосомой (фаголизосомы) либо путём слияния аутофагосомы (мембранного пузырька, содержащего собственные компоненты клетки) с первичной лизосомой (рис. 10) или поздней эндосомой (аутофаголизосомы). Аутофагия обеспечивает переваривание участков цитоплазмы, митохондрий, рибосом, фрагментов мембран и т.п. Убыль последних в клетке компенсируется их новообразованием, что ведёт к обновлению («омоложению») клеточных структур. Так, в нервных клетках человека, функционирующих многие десятилетия, большинство органоидов обновляется в течение 1 месяца.
Разновидность лизосом, содержащих непереваренные вещества (структуры), названа остаточными тельцами. Последние могут длительно находиться в цитоплазме или выделять своё содержимое путём экзоцитоза за пределы клетки (рис. 10). Распространённым видом остаточных телец в организме животных являются липофусциновые гранулы , представляющие собой мембранные пузырьки (0,3-3 мкм), содержащие труднорастворимый коричневый пигмент липофусцин.
Пероксисомы представляют собой мембранные пузырьки диаметром до 1,5 мкм, матрикс которых содержит около 15 ферментов (рис. 8). Среди последних наиболее важны каталаза, на которую приходится до 40% общего белка органоида, а также пероксидаза, оксидаза аминокислот и др. Пероксисомы образуются в эндоплазматическом ретикулуме и обновляются каждые 5-6 дней. Наряду с митохондриями, пероксисомы являются важным центром утилизации кислорода в клетке. В частности, под воздействием каталазы распадается перекись водорода (Н 2 О 2), образующаяся в ходе окисления аминокислот, углеводов и др. веществ клетки. Таким образом, пероксисомы защищают клетку от повреждающего эффекта перекиси водорода.
Органоиды энергетического обмена. Митохондрии описаны впервые Р. Келликером в 1850 году в мышцах насекомых под названием саркосом. Позднее они изучались и описывались Р. Альтманом в 1894 году как «биопласты», а в 1897 году К. Бенда назвал их митохондриями. Митохондрии представляют собой мембранные органоиды, обеспечивающие клетку (организм) энергией. Источником запасаемой в виде фосфатных связей АТФ энергии являются процессы окисления. Наряду с этим митохондрии участвуют в биосинтезе стероидов и нуклеиновых кислот, а также в окислении жирных кислот.
М Рис. 11.
Схема
строения
митохондрии:
1 – наружная
мембрана; 2 – внутренняя мембрана; 3 –
кристы; 4 – матрикс
 итохондрии
имеют эллиптическую, сферическую,
палочковидную, нитевидную и др. формы,
которые могут изменяться в течение
определенного времени. Их размеры
составляют 0,2-2 мкм в ширину и 2-10 мкм в
длину. Количество митохондрий в различных
клетках варьирует в широких пределах,
достигая в наиболее активных 500-1000. В
клетках печени (гепатоцитах) их число
составляет около 800, а занимаемый ими
объем равен примерно 20% объема цитоплазмы.
В цитоплазме митохондрии могут
располагаться диффузно, однакообычно
они сосредоточены в участках максимального
потребления энергии,
например, вблизи ионных насосов,
сократимых элементов (миофибрилл),
органелл движения (аксонема спермия).
Митохондрии состоят из
наружной и внутренней мембран,
разделенных
межмембранным пространством,
и
содержат митохондриальный матрикс, в
который обращены складки внутренней
мембраны - кристы
(рис.
11, 12).
итохондрии
имеют эллиптическую, сферическую,
палочковидную, нитевидную и др. формы,
которые могут изменяться в течение
определенного времени. Их размеры
составляют 0,2-2 мкм в ширину и 2-10 мкм в
длину. Количество митохондрий в различных
клетках варьирует в широких пределах,
достигая в наиболее активных 500-1000. В
клетках печени (гепатоцитах) их число
составляет около 800, а занимаемый ими
объем равен примерно 20% объема цитоплазмы.
В цитоплазме митохондрии могут
располагаться диффузно, однакообычно
они сосредоточены в участках максимального
потребления энергии,
например, вблизи ионных насосов,
сократимых элементов (миофибрилл),
органелл движения (аксонема спермия).
Митохондрии состоят из
наружной и внутренней мембран,
разделенных
межмембранным пространством,
и
содержат митохондриальный матрикс, в
который обращены складки внутренней
мембраны - кристы
(рис.
11, 12).
Н
Рис. 12.
Электронная
фотография митохондрии (поперечный
разрез)
Межмембранное пространство митохондрий шириной 10-20 нм содержит небольшое количество ферментов. Его ограничивает изнутри внутренняя мембрана митохондрий, содержащая транспортные белки, ферменты дыхательной цепи и сукцинатдегидрогеназу, а также комплекс АТФ-синтетазы. Внутренняя мембрана характеризуется низкой проницаемостью для мелких ионов. Она формирует складки толщиной 20 нм, которые располагаются чаще всего перпендикулярно продольной оси митохондрий, а в некоторых случаях (мышечные и др. клетки) - продольно. С повышением активности митохондрий количество складок (их общая площадь) возрастает. На кристах находятся оксисомы - грибовидные образования, состоящие из округлой головки диаметром 9 нм и ножки толщиной 3 нм. В области головки происходит синтез АТФ. Процессы окисления и синтеза АТФ в митохондриях разобщены, из-за чего не вся энергия накапливается в АТФ, рассеиваясь частично в виде тепла. Такое разобщение наиболее выражено, например, в бурой жировой ткани, используемой для весеннего «разогрева» находившихся в состоянии «зимней спячки» животных.
Внутренняя камера митохондрии (область между внутренней мембраной и кристами) заполнена матриксом (рис. 11, 12), содержащим ферменты цикла Кребса, ферменты белкового синтеза, ферменты окисления жирных кислот, митохондриальную ДНК, рибосомы и митохондриальные гранулы.
Митохондриальная ДНК представляет собственный генетический аппарат митохондрий. Она имеет вид кольцевой двухцепочечной молекулы, в которой содержится около 37 генов. Митохондриальная ДНК отличается от ядерной ДНК низким содержанием некодирующих последовательностей и отсутствием связей с гистонами. Митохондриальная ДНК кодирует иРНК, тРНК и рРНК, однако обеспечивает синтез только 5-6% митохондриальных белков (ферментов системы транспорта ионов и некоторых ферментов синтеза АТФ). Синтез всех других белков, а также удвоение митохондрий контролируются ядерной ДНК. Большая часть рибосомальных белков митохондрий синтезируется в цитоплазме, а затем транспортируется в митохондрии. Наследование митохондриальной ДНК у многих видов эукариот, включая человека, происходит только по материнской линии: митохондриальная ДНК отца исчезает при гаметогенезе и оплодотворении.
Митохондрии имеют относительно короткий жизненный цикл (около 10 суток). Разрушение их происходит путём аутофагии, а новообразование - путём деления (перешнуровки) предшествующих митохондрий. Последнему предшествует репликация митохондриальной ДНК, которая происходит независимо от репликации ядерной ДНК в любые фазы клеточного цикла.
У прокариот митохондрии отсутствуют, и их функции выполняет клеточная мембрана. Согласно одной из гипотез, митохондрии произошли из аэробных бактерий в результате симбиогенеза. Существует предположение об участии митохондрий в передаче наследственной информации.
Органоиды – постоянные и обязательные компоненты клеток; специализированные участки цитоплазмы клетки, имеющие определенную структуру и выполняющие определенные функции в клетке. Различают органоиды общего и специального назначения.
Органоиды общего назначения имеются в большинстве клеток (эндоплазматическая сеть, митохондрии, пластиды, комплекс Гольджи, лизосомы, вакуоли, клеточный центр, рибосомы). Органоиды специального назначения характерны только для специализированных клеток (миофибриллы, жгутики, реснички, сократительные и пищеварительные вакуоли). Органоиды (за исключением рибосом и клеточного центра) имеют мембранное строение.
Эндоплазматическая ретикулюм(ЭПР) – это разветвленная система соединенных между собой полостей, трубочек и каналов, образованных элементарными мембранами и пронизывающая всю толщу клетки. Открыта в 1943 г. Портером. Особенно много каналов эндоплазматической сети в клетках с интенсивным обменом веществ. В среднем объем ЭПС составляет от 30% до 50% общего объема клетки. ЭПС лабильна. Форма внутренних лакун и кана
лов, их размер, расположение в клетке и количество изменяются в процессе жизнедеятельности. Развита сильнее в животных клетка. ЭПС морфологически и функционально связана с пограничным слоем цитоплазмы, ядерной оболочкой, рибосомами, комплексом Гольджи, вакуолями, образуя вместе с ними единую функционально-структурную систему для осуществления обмена веществ и энергии и передвижения веществ внутри клетки. Вблизи эндоплазматической сети накапливаются митохондрии и пластиды.
Выделяют две разновидности ЭПС: шероховатую и гладкую. На мембранах гладкой (агранулярной) ЭПС локализованы ферменты систем жирового и углеводного синтеза: здесь происходит синтез углеводов и почти всех клеточных липидов. Мембраны гладкой разновидности эндоплазматической сети преобладают в клетках сальных желез, печени (синтез гликогена), в клетках с большим содержанием питательных веществ (семена растений). На мембране шероховатой (гранулярной) ЭПС располагаются рибосомы, где осуществляется биосинтез белков. Часть синтезируемых ими белков включается в состав мембраны эндоплазматической сети, остальные поступают в просвет ее каналов, где преобразуются и транспортируются в комплекс Гольджи. Особенно много шероховатых мембран в клетках желез и нервных клетках.
Рис. Шероховатый и гладкий эндоплазматический ретикулум.

Рис. Транспорт веществ по системе ядро – эндоплазматический ретикулум (ЭПР) – комплекс Гольджи.
Функции эндоплазматической сети :
1) синтез белков (шероховатая ЭПС), углеводов и липидов (гладкая ЭПС);
2) транспорт веществ, как поступивших в клетку, так и вновь синтезируемых;
3) деление цитоплазмы на отсеки (компартменты), что обеспечивает пространственное разделение ферментных систем, необходимое для их последовательного вступления в биохимические реакции.
Митохондрии – присутствуют практически во всех типах клеток одно- и многоклеточных организмов (за исключением эритроцитов млекопитающих). Число их в разных клетках варьирует и зависит от уровня функциональной активности клетки. В клетке печени крысы их около 2500, а в мужской половой клетке некоторых моллюсков – 20 – 22. Их больше в грудной мышце летающих птиц, чем в грудной мышце нелетающих.
Митохондрии имеют форму сферических, овальных и цилиндрических телец. Размеры составляют 0,2 – 1,0 мкм и диаметре и до 5 - 7 мкм в длину.

Рис. Митохондрия.
Длина нитевидных форм достигает 15-20 мкм. Снаружи митохондрии ограничены гладкой наружной мембраной, сходной по составу с плазмалеммой. Внутренняя мембрана образует многочисленные выросты – кристы – и содержит многочисленные ферменты, АТФ-сомы (грибовидные тела), участвующие в процессах трансформации энергии питательных веществ в энергию АТФ. Количество крист зависит от функции клетки. В митохондриях мышц крист очень много, они занимают всю внутреннюю полость органоида. В митохондриях эмбриональных клеток кристы единичны. В растительных выросты внутренней мембраны чаще имеют форму трубочек. Полость митохондрии заполнена матриксом, в котором содержатся вода, минеральные соли, белки-ферменты, аминокислоты. Митохондрии имеют автономную белоксинтезирующую систему: кольцевую молекулу ДНК, различные виды РНК и более мелкие, чем в цитоплазме рибосомы.
Митохондрии тесно связаны мембранами эндоплазматической сети, каналы которой часто открываются прямо в митохондрии. При повышении нагрузки на орган и усилении синтетических процессов, требующих затраты энергии, контакты между ЭПС и митохондриями становятся особенно многочисленными. Число митохондрий может быстро увеличиваться путем деления. Способность митохондрий к размножению обусловлена присутствием в них молекулы ДНК, напоминающей кольцевую хромосому бактерий.
Функции митохондрий :
1) синтез универсального источника энергии – АТФ;
2) синтез стероидных гормонов;
3) биосинтез специфических белков.
Пластиды – органоиды мембранного строения, характерные только для растительных клеток. В них происходят процессы синтеза углеводов, белков и жиров. По содержанию пигментов их делят на три группы: хлоропласты, хромопласты и лейкопласты.
Хлоропласты имеют относительно постоянную эллиптическую или линзовидную форму. Размер по наибольшему диаметру составляет 4 – 10 мкм. Количество в клетке колеблется от нескольких единиц до нескольких десятков. Их размер, интенсивность окраски, количество и расположение в клетке зависят от условий освещения, вида и физиологического состояния растений.

Рис. Хлоропласт, строение.
Это белково-липоидные тела, состоящие на 35-55% из белка, 20-30% - липидов, 9% - хлорофилла, 4-5% каратиноидов, 2-4% нуклеиновых кислот. Количество углеводов варьирует; обнаружено некоторое количество минеральных веществ Хлорофилл – сложный эфир органической двухосновной кислоты – хлорофиллина и органических спиртов – метилового (СН 3 ОН) и фитола (С 20 Н 39 ОН). У высших растений в хлоропластах постоянно присутствуют хлорофилл а – имеет сине-зеленую окраску, и хлорофилл b – желто-зеленую; причем содержание хлорофилла, а в несколько раз больше.
Кроме хлорофилла в состав хлоропластов входят пигменты - каротин С 40 Н 56 и ксантофилл С 40 Н 56 О 2 и некоторые другие пигменты (каратиноиды). В зеленом листе желтые спутники хлорофилла маскируются более яркой зеленой окраской. Однако осенью, при листопаде, у большинства растений хлорофилл разрушается и тогда обнаруживается присутствие в листе каратиноида – лист становится желтым.
Хлоропласт одет двойной оболочкой, состоящей из наружной и внутренней мембран. Внутреннее содержимое – строма – имеет ламеллярное (пластинчатое) строение. В бесцветной строме выделяют граны – окрашенные в зеленые цвет тельца, 0,3 – 1,7 мкм. Они представляют собой совокупность тилакоидов – замкнутых телец в виде плоских пузырьков или дисков мембранного происхождения. Хлорофилл в виде мономолекулярного слоя располагается между белковым и липидным слоями в тесной связи с ними. Пространственное расположение молекул пигментов в мембранных структурах хлоропластов является весьма целесообразным и создает оптимальные условия для наиболее эффективного поглощения, передачи и использования лучистой энергии. Липиды образуют безводные диэлектрические слои мембран хлоропласта, необходимые для функционирования электронно-транспортной цепи. Роль звеньев цепи переноса электронов выполняют белки (цитохромы, пластохиноны, ферредоксин, пластоцианин) и отдельные химические элементы – железо, марганец и др. Количество гран в хлоропласте от 20 до 200. Между гранами, связывая их друг с другом, располагаются ламеллы стромы. Ламеллы гран и ламеллы стромы имеют мембранное строение.
Внутренне строение хлоропласта делает возможным пространственное разобщение многочисленных и разнообразных реакций, составляющих в своей совокупности содержание фотосинтеза.
Хлоропласты, как и митохондрии, содержат специфическую РНК и ДНК, а также более мелкие рибосомы и весь молекулярный арсенал, необходимый для биосинтеза белка. У этих органоидов имеется достаточное для обеспечения максимальной активности белоксинтезирующей системы количество и-РНК. Вместе с тем в них содержится и достаточно ДНК для кодирования определенных белков. Они размножаются делением, путем простой перетяжки.
Установлено, что хлоропласты могут изменять свою форму, размеры и положение в клетке, т. е. способны самостоятельно двигаться (таксис хлоропластов). В них обнаружено два типа сократительных белков, за счет которых, очевидно, и осуществляется активное движение этих органоидов в цитоплазме.
Хромопласты широко распространены в генеративных органах растений. Они окрашивают лепестки цветков (лютика, георгина, подсолнечника), плоды (томатов, рябины, шиповника) в желтый, оранжевый, красный цвета. В вегетативных органах хромопласты встречаются значительно реже.
Окраска хромопластов обусловлена присутствием каратиноидов – каротина, ксантофилла и ликопина, которые в пластидах находятся в различном состоянии: в виде кристаллов, липоидного раствора или в соединении с белками.
Хромопласты, по сравнению с хлоропластами, имеют более простое строение – в них отсутствует ламеллярная структура. Химический состав также отличен: пигменты – 20–50%, липиды до 50%, белки – около 20%, РНК – 2-3%. Это свидетельствует о меньшей физиологической активности хлоропластов.
Лейкопласты не содержат пигментов, они бесцветны. Эти самые мелкие пластиды имеют округлую, яйцевидную или палочковидную форму. В клетке они часто группируются вокруг ядра.
Внутренне структура, еще менее дифференцирована по сравнению с хлоропластами. В них осуществляется синтез крахмала, жиров, белков. В соответствии с этим выделяют три вида лейкопластов – амилопласты (крахмал), олеопласты (растительные масла) и протеопласты (белки).
Возникают лейкопласты из пропластид, с которыми они сходны по форме и строению, а отличаются лишь размерами.
Все пластиды генетически связаны друг с другом. Они образуются из пропластид – мельчайших бесцветных цитоплазматических образований, сходных по внешнему виду с митохондриями. Пропластиды находятся в спорах, яйцеклетках, в эмбриональных клетках точек роста. Непосредственно из пропластид образуются хлоропласты (на свету) и лейкопласты (в темноте), а из них развиваются хромопласты, являющиеся конечным продуктом в эволюции пластид в клетке.

Комплекс Гольджи – впервые был обнаружен в 1898 г. Итальянским ученым Гольджи в животных клетках. Это система внутренних полостей, цистерн (5-20), располагающихся сближено и параллельно друг другу, и крупных и мелких вакуолей. Все эти образования имеют мембранное строение и являются специализированными участками эндоплазматической сети. В животных клетках комплекс Гольджи развит лучше, чем в растительных; в последних он называется диктиосомы.

Рис. Строение комплекса Гольджи.
Попадающие в пластинчатый комплекс белки и липиды, подвергаются различным преобразованиям, накапливаются, сортируются, упаковываются в секреторные пузырьки и транспортируются по назначению: к различным структурам внутри клетки или за пределы клетки. Мембраны комплекса Гольджи также синтезируют полисахариды и образуют лизосомы. В клетках молочных желез комплекс Гольджи участвует в образовании молока, а в клетках печени – желчи.
Функции комплекса Гольджи :
1) концентрация, обезвоживание и уплотнение синтезированных в клетке белков, жиров, полисахаридов и веществ, поступивших извне;
2) сборка сложных комплексов органических веществ и подготовка их к выведению из клетки (целлюлоза и гемицеллюлоза у растений, гликопротеины и гликолипиды у животных);
3) синтез полисахаридов;
4) образование первичных лизосом.
Лизосомы - небольшие овальные тельца диаметром 0,2-2,0 мкм. Центральное положение занимает вакуоль, содержащая 40 (по разным данным 30-60) гидролитических ферментов, способных в кислой среде (рН 4,5-5) расщеплять белки, нуклеиновые кислоты, полисахариды, липиды и другие вещества.
Вокруг этой полости располагается строма, одетая снаружи элементарной мембраной. Расщепление веществ при помощи ферментов называется лизисом, поэтому органоид назван лизосомой. Образование лизосом происходит в комплексе Гольджи. Первичные лизосомы приближаются непосредственно к пиноцитозным или фагоцитозным вакуолям (эндосомам) и изливают свое содержимое в их полость, образуя вторичные лизосомы (фагосомы), внутри которых и происходит переваривание веществ. Продукты лизиса через мембрану лизосом поступают в цитоплазму и включаются в дальнейший обмен веществ. Вторичные лизосомы с остатками непереваренных веществ называются остаточными тельцами. Примером вторичных лизосом являются пищеварительные вакуоли простейших.
Функции лизосом :
1) внутриклеточное переваривание макромолекул пищи и чужеродных компонентов, поступающих в клетку при пино- и фагоцитозе, обеспечивая клетку дополнительным сырьем для биохимических и энергетических процессов;
2) при голодании лизосомы переваривают некоторые органоиды и на какое-то время пополняют запас питательных веществ;
3) разрушение временных органов эмбрионов и личинок (хвост и жабры у лягушки) в процессе постэмбрионального развития;

Рис. Образование лизосом
Вакуоли – полости в цитоплазме растительных клеток и протист, заполненные жидкостью. Имеют форму пузырьков, тонких канальцев и другую. Вакуоли образуются из расширений эндоплазматической сети и пузырьков комплекса Гольджи как тончайшие полости, затем по мере роста клетки и накопления продуктов обмена объем их увеличивается, а количество сокращается. Развитая сформировавшаяся клетка имеет обычно одну большую вакуоль, занимающую центральное положение.
Вакуоли растительных клеток заполнены клеточным соком, который представляет собой водный раствор органических (яблочная, щавелевая, лимонная кислоты, сахара, инулин, аминокислоты, белки, дубильные вещества, алкалоиды, глюкозиды) и минеральных (нитраты, хлориды, фосфаты) веществ.
У протист встречаются пищеварительные вакуоли и сократительные.
Функции вакуолей :
1) хранилища запасных питательных веществ и вместилища выделений (у растений);
2) определяют и поддерживают осмотическое давление в клетках;
3) обеспечивают внутриклеточное пищеварение у протист.

Рис. Клеточный центр.
Клеточный центр обычно находится вблизи ядра и состоит из двух центриолей, расположенных перпендикулярно друг другу и окруженных лучистой сферой. Каждая центриоль представляет собой полое цилиндрическое тельце длиной 0,3-0,5 мкм и длиной 0,15 мкм, стенка которого образована 9 триплетами микротрубочек. Если центриоль лежит в основании реснички или жгутика, то ее называют базальным тельцем .
Перед делением центриоли расходятся к противоположным полюсам и возле каждой из них возникает дочерняя центриоль. От центриолей, расположенных на разных полюсах клетки, образуются микротрубочки, растущие навстречу друг другу. Они формируют митотическое веретено, способствующее равномерному распределению генетического материала между дочерними клетками, являются центром организации цитоскелета. Часть нитей веретена прикрепляется к хромосомам. В клетках высших растений клеточный центр центриолей не имеет.
Центриоли относятся к самовоспроизводящимся органоидам цитоплазмы. Они возникают в результате дупликации уже имеющихся. Это происходит при расхождении центриолей. Незрелая центриоль содержит 9 одиночных микротрубочек; по-видимому, каждая микротрубочка является матрицей при сборке триплетов, характерных для зрелой центриоли.
Центросома характерна для клеток животных, некоторых грибов, водорослей, мхов и папоротников.
Функции клеточного центра :
1) образование полюсов деления и формирование микротрубочек веретена деления.
Рибосомы - мелкие сферические органоиды, от 15 до 35 нм. Состоят из двух субъединиц большой (60S) и малой (40S). Содержат около 60% белка и 40% рибосомальной РНК. Молекулы рРНК образуют ее структурный каркас. Большинство белков специфически связано с определенными участками рРНК. Некоторые белки входят в состав рибосом только во время биосинтеза белка. Субъединицы рибосом образуются в ядрышках. и через поры в ядерной оболочке поступают в цитоплазму, где располагаются либо на мембране ЭПА, либо на наружной стороне ядерной оболочки, либо свободно в цитоплазме. Сначала на ядрышковой ДНК синтезируются рРНК, которые затем покрываются поступающими из цитоплазмы рибосомальными белками, расщепляются до нужных размеров и формируют субъединицы рибосом. Полностью сформированных рибосом в ядре нет. Объединение субъединиц в целую рибосому происходит в цитоплазме, как правило, во время биосинтеза белка. По сравнению с митохондриями, пластидами, клетками прокариот рибосомы в цитоплазме эукариотических клеток крупнее. Могут объединяться по 5-70 единиц в полисомы.
Функции рибосом :
1) участие в биосинтезе белка.

Рис. 287. Рибосома: 1 - малая субъединица; 2 - большая субъединица.
Реснички, жгутики – выросты цитоплазмы, покрытые элементарной мембраной, под которой находится 20 микротрубочек, образующих 9 пар по периферии и две одиночные в центре. У основания ресничек и жгутиков расположены базальные тельца. Длина жгутиков достигает 100 мкм. Реснички – это короткие – 10-20 мкм – жгутики. Движение жгутиков винтовое, а ресничек – веслообразное. Благодаря ресничкам и жгутикам передвигаются бактерии, протисты, ресничные, перемещаются частицы или жидкости (реснички мерцательного эпителия дыхательных путей, яйцеводов), половые клетки (сперматозоиды).

Рис. Строение жгутиков и ресничек эукариот
Включения - временные компоненты цитоплазмы, то возникающие, то исчезающие. Как правило, они содержатся в клетках на определенных этапах жизненного цикла. Специфика включений зависит от специфики соответствующих клеток тканей и органов. Включения встречаются преимущественно в растительных клетках. Они могут возникать в гиалоплазме, различных органеллах, реже в клеточной стенке.

В функциональном отношении включения представляют собой либо временно выведенные из обмена веществ клетки соединения (запасные вещества - крахмальные зерна, липидные капли и отложения белков), либо конечные продукты обмена (кристаллы некоторых веществ).
Крахмальные зерна . Это наиболее распространенные включения растительных клеток. Крахмал запасается у растений исключительно в виде крахмальных зерен. Они образуются только в строме пластид живых клеток. В процессе фотосинтеза в зеленых листьях образуется ассимиляционный , или первичный крахмал. Ассимиляционный крахмал в листьях не накапливается и, быстро гидролизуясь до сахаров, оттекает в части растения, в которых происходит его накопление. Там он вновь превращается в крахмал, который называют вторичным. Вторичный крахмал образуется и непосредственно в клубнях, корневищах, семенах, то есть там, где он откладывается в запас. Тогда его называют запасным . Лейкопласты, накапливающие крахмал, называют амилопластами . Особенно богаты крахмалом семена, подземные побеги (клубни, луковицы, корневища), паренхима проводящих тканей корней и стеблей древесных растений.
Липидные капли . Встречаются практически во всех растительных клетках. Наиболее богаты ими семена и плоды. Жирные масла в виде липидных капель - вторая по значению (после крахмала) форма запасных питательных веществ. Семена некоторых растений (подсолнечник, хлопчатник и т.д.) могут накапливать до 40% масла от массы сухого вещества.
Липидные капли, как правило, накапливаются непосредственно в гиалоплазме. Они представляют собой сферические тела обычно субмикроскопического размера. Липидные капли могут накапливаться и в лейкопластах, которые называют элайопластами .
Белковые включения образуются в различных органеллах клетки в виде аморфных или кристаллических отложений разнообразной формы и строения. Наиболее часто кристаллы можно встретить в ядре - в нуклеоплазме, иногда в перинуклеарном пространстве, реже в гиалоплазме, строме пластид, в расширениях цистерн ЭПР, матриксе пероксисом и митохондриях. В вакуолях встречаются как кристаллические, так и аморфные белковые включения. В наибольшем количестве кристаллы белка встречаются в запасающих клетках сухих семян в виде так называемых алейроновых 3 зерен или белковых телец .
Запасные белки синтезируются рибосомами во время развития семени и откладываются в вакуоли. При созревании семян, сопровождающемся их обезвоживанием, белковые вакуоли высыхают, и белок кристаллизуется. В результате этого в зрелом сухом семени белковые вакуоли превращаются в белковые тельца (алейроновые зерна).
Статьи по теме